
Abstract
Despite being known from every continent, the geological record of pterosaurs, the first group of vertebrates to develop powered flight, is very uneven, with only a few deposits accounting for the vast majority of specimens and almost half of the taxonomic diversity. Among the regions that stand out for the greatest gaps of knowledge regarding these flying reptiles, is the Afro-Arabian continent, which has yielded only a small number of very fragmentary and incomplete materials. Here we fill part of that gap and report on the most complete pterosaur recovered from this continent, more specifically from the Late Cretaceous (~95 mya) Hjoûla Lagerstätte of Lebanon. This deposit is known since the Middle Ages for the exquisitely preserved fishes and invertebrates, but not for tetrapods, which are exceedingly rare. Mimodactylus libanensis gen. et sp. nov. differs from the other Afro-Arabian pterosaur species named to date and is closely related to the Chinese species Haopterus gracilis, forming a new clade of derived toothed pterosaurs. Mimodactylidae clade nov. groups species that are related to Istiodactylidae, jointly designated as Istiodactyliformes (clade nov.). Istiodactyliforms were previously documented only in Early Cretaceous sites from Europe and Asia, with Mimodactylus libanensis the first record in Gondwana.
Introduction
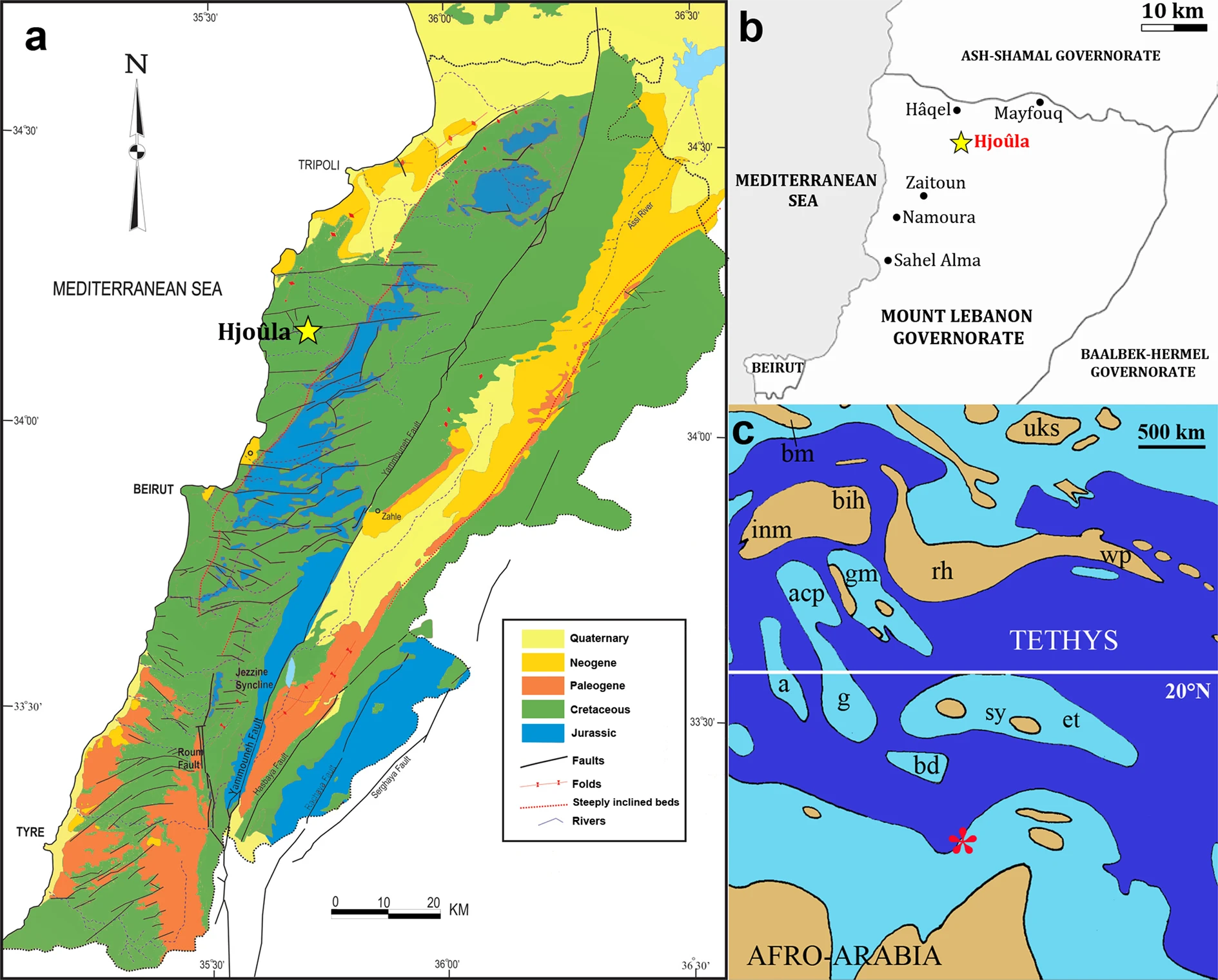
Concerning Mesozoic vertebrate palaeontology, the Afro-Arabian continent is still veiled in mystery. Except for South Africa, where systematic studies, mainly in Triassic and Lower Jurassic continental deposits have provided a comparatively diverse vertebrate fauna1, the information about the biota that lived during most of the Mesozoic Era in this region is extremely limited. This is particularly true for pterosaurs, an extinct group of flying reptiles that includes the major powered flying vertebrates for almost 160 mya2,3. The main African records of this group are restricted to isolated elements from the Jurassic deposits of Tendaguru4,5,6 from Tanzania, and the Upper Cretaceous Kem Kem Beds (Cenomanian)7,8,9,10,11 and Ouled Abdoun (Maastrichtian)12,13 from Morocco. The most complete pterosaur specimens from the Afro-Arabian continent have been recovered from Cenomanian marine deposits of Lebanon14 (Fig. 1). The first specimen was a partial left forelimb of a relatively small unnamed ornithocheiroid from Hâqel Lagerstätte15 and the second a crushed skeleton formed mainly by two wings and the shoulder girdle of the azhdarchoid Microtuban altivolans16 from the coeval14,17 Hjoûla Lagerstätte.
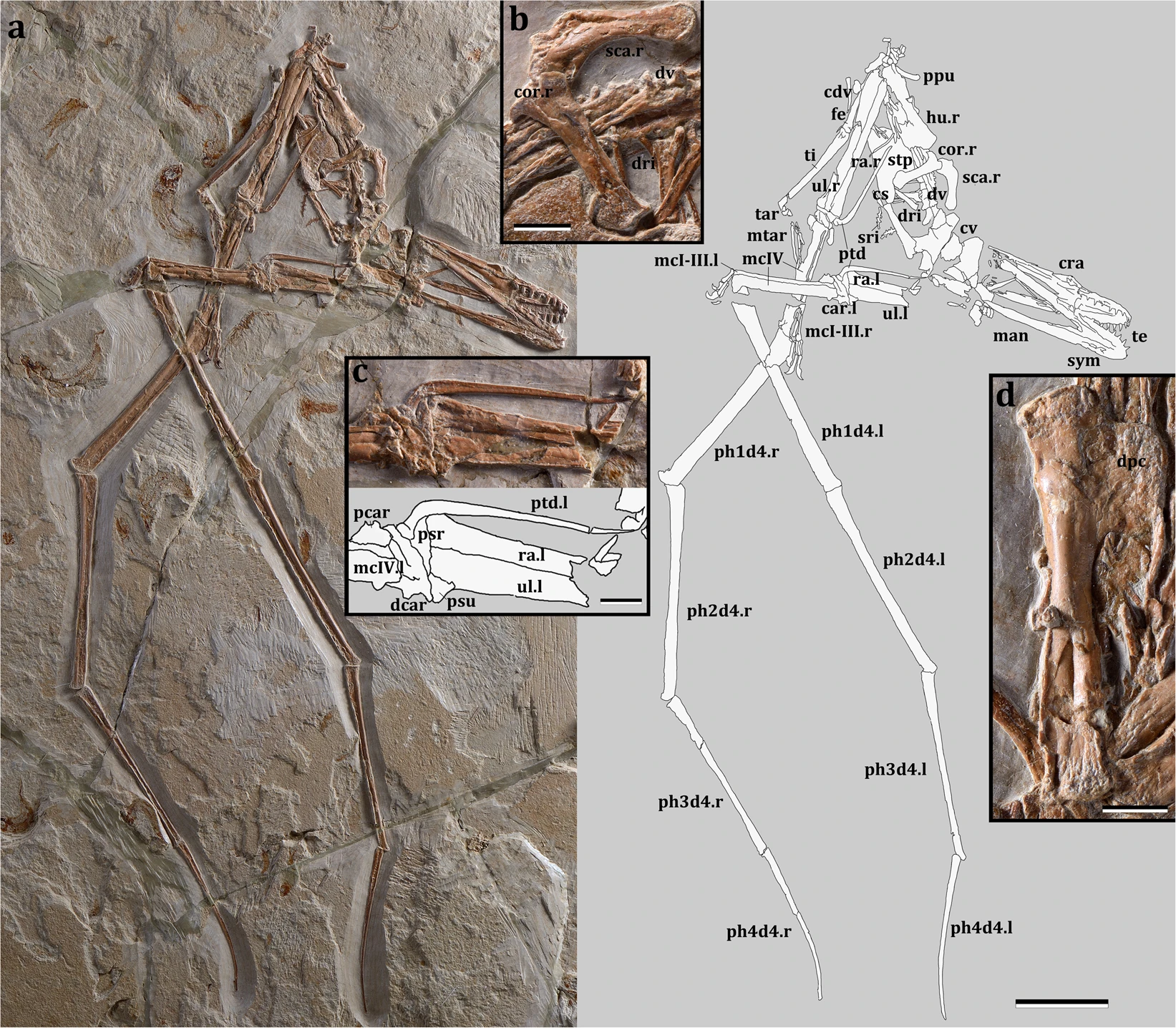
The specimen described here (Fig. 2) is the first complete and articulated skeleton including the skull and lower jaw from the Afro-Arabian continent providing new insights on the diversity and potential palaeoecology of ornithocheiroids.
Results
Systematic palaeontology
Pterosauria Kaup, 1834.
Pterodactyloidea Plieninger, 1901.
Ornithocheiroidea Seeley, 1870 sensu Kellner (2003)18.
Pteranodontoidea Marsh, 1876 sensu Kellner18.
Lanceodontia Andres et al.19.
Istiodactyliformes clade nov.
Branch-based definition
The most inclusive clade containing Istiodactylus latidens, but not Anhanguera blittersdorffi.
Diagnosis
Slender-built lanceodontian pterodactyloids with the following synapomorphies: mandibular rostral end pointed, teeth confined to the anterior half of the jaws, and labiolingually compressed crowns with a cingulum.
Included taxa
Istiodactylidae, Mimodactylidae, and Hongshanopterus lacustris.
Mimodactylidae clade nov.
Branch-based definition
The most inclusive clade containing Mimodactylus libanensis gen. et sp. nov., but not Istiodactylus latidens, Ikrandraco avatar, and Anhanguera blittersdorffi.
Diagnosis
Istiodactyliforms with cone-shaped teeth, crowns with a slight labiolingual compression, and sternal articular surface of the coracoid slightly concave.
Included species
Haopterus gracilis and Mimodactylus libanensis gen. et sp. nov.
Mimodactylus libanensis gen. et sp. nov.
Etymology
Mimodactylus, from the acronym MIM (Mineral Museum) for the museum housing the specimen, in recognition of both the museum and the wishes of the anonymous philanthropist who facilitated the acquisition of the specimen thus keeping it in Lebanon, and the Greek ‘dactylos’ (δάκτυλος), meaning digit; libanensis from Lebanon, where the specimen was found.
Holotype
Almost complete skeleton, including the skull and lower jaw, housed at the Mineral Museum (MIM) of Beirut, Lebanon, MIM F1. Cast at the University of Alberta, Edmonton and Museu Nacional/UFRJ (MN 7216-V).
Locality and horizon
Hjoûla Lagerstätte of the Sannine Limestone (late Cenomanian17), near the town of Hjoûla located 35 km NNE of Beirut and 10 km inland from Jbail, Lebanon.
Diagnosis
Mimodactylid with the following autapomorphies: humerus with a rectangular deltopectoral crest; humerus smaller than half the length of the second phalanx of the wing finger (hu/ph2d4 < 0.5). This species can be further distinguished from other ornithocheiroid pterodactyloids by the following combination of characters: discrete palatal ridge; 11 and 10 cone-shaped teeth on each side of the upper and lower jaws, respectively; scapula slightly longer than coracoid; humerus much longer than femur (hu/fe ~1.3); deltopectoral crest extends for around 40% of the humerus shaft length (see Supplementary Information for further details and measurements).
Comparative description
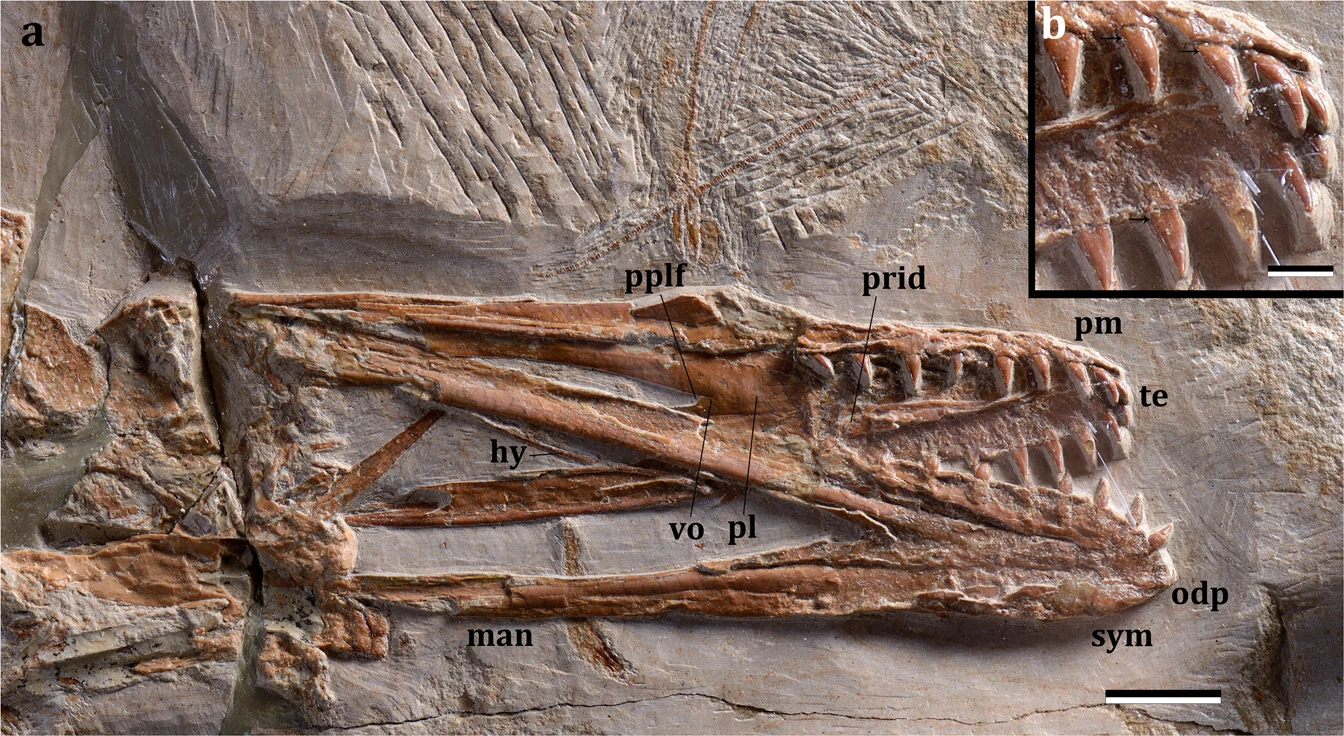
The specimen is well preserved with most parts of the skeleton articulated or only slightly displaced from their anatomical position (Fig. 2a). The skull and lower jaw are exposed in ventral view, with the occipital region and the craniomandibular articulation flattened. It is a comparatively small individual, with a wingspan of ~1.32 meters, and long wings, resulting in a high aspect ratio. Based on the unfused scapula and coracoid, pelvic elements and sacral vertebrae but fused dentaries at the symphysis, and dorsal vertebrae not fused into a notarium, it is likely that it was a very young animal at the time of death, having reached an ontogenetic stage between 2 and 320. The skull has a broad rostrum in dorsopalatal view (Fig. 3), but not as rounded as in Istiodactylus21,22,23 and also differing from other istiodactyliforms24,25. The rostral tip is pointed, unlike the rounded terminus of Istiodactylus21,22,23. There are 11 and 10 cone-shaped teeth on each side of the upper and lower jaws, respectively (Fig. 3a), similar to Haopterus26 and Linlongopterus27. Crowns are labiolingually compressed with a cingulum (Fig. 3b; SI) as in Haopterus and other istiodactyliforms. This cingulum was previously reported in Istiodactylidae28 and related taxa, but the teeth of this latter lineage is characterised by wide crowns, which have also a marked labiolingually compression21,22,23,24,25. The palate is concave and shows a small palatal ridge. Choanae are large and divided by the vomers. The postpalatinal fenestra has an elongated egg-like shape as in the basal istiodactyliform Hongshanopterus25. Ceratobranchials I of the hyoid apparatus are fork-like, thin, and elongate elements. An odontoid process is present at the tip of the lower jaw as in Istiodactylus latidens23, but this process could also be recognised in Haopterus26 and Lonchodraco giganteus29. The dorsal vertebrae (Fig. 2b; Supplementary Fig. S1) are exposed in ventral view and are not fused into a notarium. A total of 7 caudal vertebrae were identified (Supplementary Fig. S2), all of which lack a duplex centrum and decrease rapidly in size posteriorly, suggesting that this species had a short tail. The cristospine of the sternum is comparatively short and deep, similar to those of Nurhachius and Istiodactylus; the anterior portion of the sternum is more rounded in lateral view than that of istiodactylids, being, in this respect, more similar to that of the Anhangueridae. The scapula is stout and shares with istiodactylids and anhanguerids a constricted shaft (Fig. 2b). However, it differs from both by being longer than the coracoid18. The coracoid sternal articulation is slightly concave as in Haopterus25 and has a developed posterior expansion that is not present in istiodactylids. The humerus (Fig. 2d) has a rectangular deltopectoral crest with an unusual straight distal margin and extends approximately 40% down the humerus shaft, more than in any other ornithocheiroid except for Pteranodon and related taxa30. Mimodactylus has some wing elements longer relative to the humerus compared to istiodactylids, in particular the first and second phalanges. The distal portion of the last phalanx of the wing finger is curved as in most pterosaurs. The feet are relatively small, similar to istiodactylids30. The pteroid (Fig. 2c) is quite large (longer than the humerus). This bone is clearly articulated with the proximal syncarpal and directed towards the body. There has been a long discussion about the position of this unique pterosaur bone with the carpal elements31,32,33, which is clearly settled in the present specimen whose forelimb bones are perfectly articulated.
Discussion
Despite the incompleteness of the two other pterosaur specimens described from the upper Cenomanian of Lebanon, both of which lack cranial elements, those specimens can clearly be distinguished from Mimodactylus. The only complete bones of the ornithocheiroid MSNM V 38818 are the wing metacarpal and the radius + ulna, whose proportions differ from those of Mimodactylus showing that the latter has a proportionally longer forearm. Furthermore, the diameter of the radius of MSNM V 3881 is less than half that of the ulna, contrary to the condition of Mimodactylus. The second specimen, the holotype of Microtuban altivolans16, has a much shorter wing, a humerus with a different deltopectoral crest and a scapula that lacks the constricted shaft observed in Mimodactylus.
Another interesting feature of Mimodactylus is the dentition (Figs. 3a and SI), which differs from that of most ornithocheroids. As in Haopterus25 and Linlongopterus26, the new species has cone-shaped dental crowns and they are confined to the anterior half part of the jaws. Such a configuration is present in other istiodactyliforms and cannot be established in Lonchodraco giganteus due to preservation29, which also present cone-shaped teeth. As in Haopterus but unlike Linlongopterus and Lonchodraco, teeth are characterised by a cingulum at the base of the crown (Supplementary Fig. S3), which is also present in the Istiodactylidae and closely related species. Mimodactylus, however, lacks the lancet-shaped teeth with marked labiolingually compressed crowns that are diagnostic of the istiodactylids20,21,22,23,24. The new species also lacks the sharp carinae reported in Istiodactylus21. The first upper tooth of Mimodactylus is comparatively small and has a sub-circular transverse section. It is followed by the largest teeth in the upper jaw, which have slight labiolingually compressed crowns with a cingulum, convex labial surfaces and thin, lingually inclined, needle-like tips. This general morphology is present in the remaining teeth, also from the lower jaw. This kind of dentition is more similar to that of the basal archaeopterodactyloids18 Pterodactylus and Germanodactylus rhamphastinus24 than to istiodactylids and ornithocheiroids. The sole other derived pterodactyloid with a comparable dentition is Haopterus gracilis, first regarded as an archaeopterodactyloid26, later as an ornithocheroid close to Istiodactylidae28, and even the sister taxa of Ornithocheiroidea13,19. Here we recovered Haopterus at the base of Laceodontia as in more recent phylogenetic analyses34,35, forming a clade with Mimodactylus libanensis (Fig. 4a; see Supplementary Information for further details).
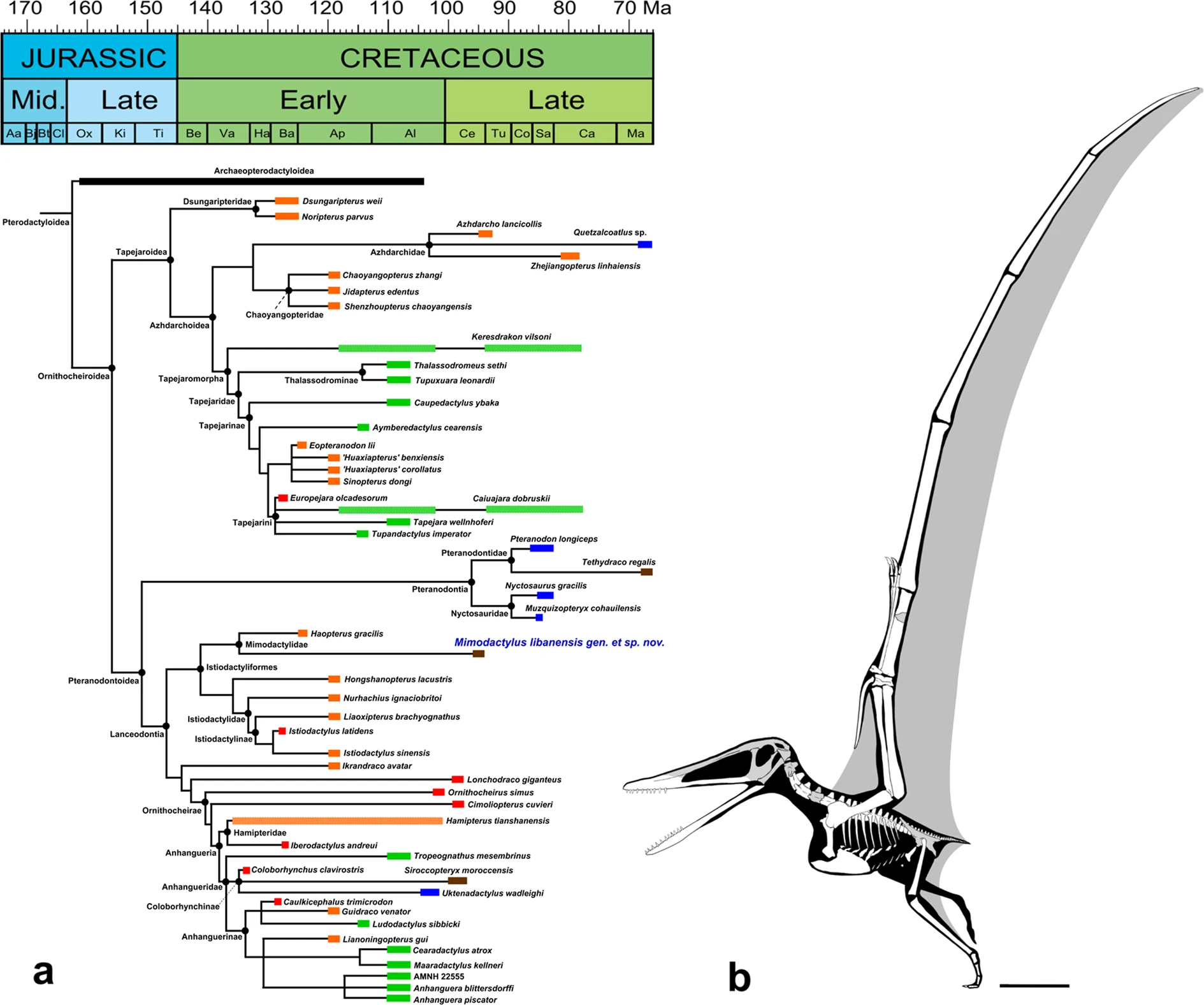
The Mimodactylidae clade nov. is diagnosed by the following synapomorphies: cone-shaped teeth on each side of the upper jaws, crowns with a slight labiolingual compression, and sternal articular surface of the coracoid slightly concave. In addition, all mimodactylids have the teeth confined to the anterior half of the jaws and are widely spaced. Although several characters presented previously are recorded in mimodactylids and other istiodactyliforms, some of them are recovered as symplesiomorphies shared by other lanceodonts (e.g., character state 77(1): presence of an odontoid process in the lower jaw; character state 95(1): cone-shaped teeth) or are of unclear origin (e.g., character state 15(3): elongated egg-shaped postpalatine fenestra; character state 56(3): quadrate inclined about 150° posteriorly to ventral margin of the skull). In addition, two synapomorphies that support the Mimodactylidae should be considered with caution due to the missing data in most of lanceodontians30 (see Supplementary Information for further details) and the potential of ontogenetic variation in derived pterodactyloids18,36.
Despite the inherent difficulties of establishing the diet of extinct vertebrates with no suitable modern analogues such as pterosaurs, the following feeding habits have been proposed for derived pterodactyloids37, mainly based on their dentition (or absence of teeth) and the shape of their rostra: piscivory for Anhangueridae and their kin30,38,39,40, Ikrandraco41, Pteranodontidae30, Nyctosauridae42, Chaoyangopteridae43, and Thalassodrominae44; frugivory for Tapejarinae45,46; durophagy for Dsungaripterus and related species30; insectivory for Nemicolopterus47; scavenging for Istiodactylus21,22; cutting or ploughing through unconsolidated sediments for Argentinadraco48, and terrestrial stalking for the long-necked azhdarchids49. The dentition of Mimodactylus differs from all of them, suggesting that this lineage of derived pterodactyloid had a different feeding habit.
Studies on the shapes of teeth of extant insectivorous tetrapods emphasise that insectivorous species profit from having slimmer teeth that could be more easily used to breakdown arthropods due to the slight radius of curvature50,51,52,53. Within the pterosaur spectrum, the anurognathids that have well-spaced and isodont teeth have been regarded as insectivores25,54. Although wider, the tooth structure of Mimodactylus may suggest a similar feeding habit, allowing them to break up arthropod exoskeletons.
Aerial insectivory is closely linked to the ability to maneuver during flight55,56,57. Extant vertebrate aerial insectivores exhibit short wings with low aspect ratios that allow them to be highly maneuverable in the air56,57,58, contrary to Mimodactylus libanensis, which had long wings with high aspect ratio (Fig. 4b). In Mimodactylus, as open-sea flyers, the ability to maneuver during flight appears to be limited and it was likely high stable during flight as observed in albatrosses and other birds (Fig. 5). This might also have been the case for some large pterosaurs such as anhanguerians, istiodactylids and pteranodontians, which are considered to have conducted a dynamic soaring58. Therefore, alternately to the insectivore hypothesis, Mimodactylus and their relatives might also have been capable of foraging for decapod crustaceans on surface waters, just like some albatross species feed upon caridean or penaeid shrimps59. In addition, a broad rostrum60 and spaced but relatively robust and pointed teeth61,62 could be good tools to seize shrimps in the water.

Observing the fossil content of the Hjoûla Lagerstätte as well as of other Cretaceous Lebanese Lagerstätten, no insects were recovered so far63,64. Even terrestrial plants are extremely rare at Hjoûla65, suggesting that this Lebanese Lagerstätte was far away from emergent areas such as islands, with the continent several hundred kilometres away. On the other hand, decapod crustaceans are the most common invertebrates found in Hjoûla66,67. No taphonomic bias was detected to explain the absence of insects, pointing to fishes and zooplankton as potentially the main local source of food for pterosaurs. Mimodactylus libanensis also has a broad rostrum, which is consistent with a faunivorous feeding habit – or primarily feeding on crustaceans – as present in extant ducks, boat-billed herons, and shoebills68. Although insectivory cannot be ruled out, all available evidence suggests that Mimodactylus was feeding on crustaceans.
This new pterosaur lived in archipelagos and scattered islands, which were present during the Late Cretaceous in the gigantic carbonate platform bordering the northern part of the Afro-Arabian continent with the Neotethys (Fig. 5). The discovery of Mimodactylus libanensis expands the spectrum of possible feeding strategies in derived pterodactyloids, a group of fascinating volant reptiles for which we still know very little.
Methods
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature. The ZooBank Life Science Identifiers (LSIDs) can be resolved and the associated information viewed by appending the LSIDs to the prefix http://zoobank.org/. The LSID for this publication is urn:lsid:zoobank.org:pub:E77F30DB-D268-4C96-9849-1B65BCDDCAA3:, and the LSIDs for the new erected groups and taxa are: urn:lsid:zoobank.org:act:E33BF241-AD34-4878-AE3D-45F13A97F327 (Istiodactyliformes), urn:lsid:zoobank.org:act:A482B812-EA6A-4EDA-8986-3EFF1D7451B2 (Mimodactylidae), urn:lsid:zoobank.org:act:28DC7F4E-C3C3-4459-B3F1-FF58BBEF3A66 (Mimodactylus), and urn:lsid:zoobank.org:act:DCD7BFC3-0F35-4AA3-91B4-89A7604EAEB7 (Mimodactylus libanensis).
Phylogenetic analysis
We performed a phylogenetic analysis using the software TNT 1.569. This analysis is based essentially on Holgado et al.34 (for further details see Supplementary Information). Search for the most parsimonious trees (MPTs) was conducted via Traditional Search (TBR swapping algorithm), 10,000 replicates, random seed and collapsing trees after search.
References
- 1.
Durand, J. F. African contributions to Paleozoic and Mesozoic vertebrate Paleontology. Jour. African Earth Sci. 43, 53–82 (2005).
- 2.
Barrett, P. M., Butler, R. J., Edwards, N. P. & Milner, A. R. Pterosaur distribution in time and space: an atlas. Zitteliana 28, 61–107 (2008).
- 3.
Hone, D. W. E. Pterosaur Research: Recent Advances and a Future Revolution. Acta Geol. Sin. 86, 1366–1376 (2012).
- 4.
Reck, H. Die deutschostafrikanischen Flugsaurier. Zentralblatt für Mineralogie, Geologie und Paläontologie. Abt. B 7, 321–336 (1931).
- 5.
Unwin, D. M. & Heinrich, W. D. On a pterosaur jaw from the Upper Jurassic of Tendaguru (Tanzania). Mitt. Mus. Nat.kd. Berl., Geowiss.Reihe 2, 121–134 (1999).
- 6.
Costa, F. R., Sayão, J. M. & Kellner, A. W. A. New pterosaur material from the Upper Jurassic of Tendaguru (Tanzania) Africa. Hist. Biol. 27, 646–655 (2015).
- 7.
Mader, B. J. & Kellner, A. W. A. A new anhanguerid pterosaur from the Cretaceous of Morocco. Bol. Museu Nac. 45, 1–11 (1999).
- 8.
Kellner, A. W. A., Mello, A. M. S. & Ford, T. A survey of pterosaurs from Africa with the description of a new specimen from Morocco. In: Carvalho, I. S. et al. (Eds). Paleontologia: Cenários da Vida, Vol. 1. Interciência 257–267 (2007).
- 9.
Rodrigues, T., Kellner, A. W. A., Mader, B. J. & Russell, D. A. New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco. Riv. Ital. Paleontol. S. 117(1), 149–160 (2011).
- 10.
Martill, D. M., Unwin, D. M., Ibrahim, N. & Longrich, N. A new edentulous pterosaur from the Cretaceous Kem Kem beds of south eastern Morocco. Cretac. Res. 84, 1–12 (2018).
- 11.
Jacobs, M. L., Martill, D. M., Ibrahim, N. & Longrich, N. A new species of Coloborhynchus (Pterosauria, Ornithocheiridae) from the mid-Cretaceous of North Africa. Cretac. Res. 95, 77–88 (2019).
- 12.
Pereda-Suberbiola, X. et al. A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geol. Soc. London Spec. Publ. 217(1), 79–90 (2003).
- 13.
Longrich, N. R., Martill, D. M. & Andres, B. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLoS Biol 16(3), e2001663 (2018).
- 14.
Dubertret, L. Carte géologique du Liban au 200 000e. Ministère des Travaux publics, Beyrouth (1955).
- 15.
Dalla Vecchia, F. M., Arduini, P. & Kellner, A. W. A. The first pterosaur from the Cenomanian (Late Cretaceous) Lagerstätten of Lebanon. Cretac. Res. 22, 219–225 (2001).
- 16.
Elgin, R. A. & Frey, E. A new azhdarchoid pterosaur from the Cenomanian (late Cretaceous) of Lebanon. Swiss J. Geosci. 104, 21–33 (2011).
- 17.
Wippich, M. G. E. & Lehmann, J. Allocrioceras from the Cenomanian (Mid-Cretaceous) of Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites. Palaeontology 47, 1093–1107 (2004).
- 18.
Kellner, A. W. A. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol. Soc. London Spec. Publ. 217(1), 105–137 (2003).
- 19.
Andres, B., Clark, J. & Xu, X. The Earliest Pterodactyloid and the Origin of the Group. Curr. Biol. 24, 1011–1016 (2014).
- 20.
Kellner, A. W. A. Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa. An. Acad. Bras. Ciênc. 87, 669–689 (2015).
- 21.
Andres, B. & Qiang, J. Istiodactylus (Pterosauria, Pterodactyloidea) from the Lower Cretaceous of Liaoning, China. J. Vert. Paleontol. 26, 70–78 (2006).
- 22.
Witton, M. P. New Insights into the Skull of Istiodactylus latidens (Ornithocheiroidea, Pterodactyloidea). PLoS One 7, e33170 (2012).
- 23.
Martill, D. M. A functional odontoid in the dentary of the Early Cretaceous pterosaur Istiodactylus latidens: Implications for feeding. Cretac. Res. 47, 56–65 (2014).
- 24.
Wang, X., Kellner, A. W. A., Zhou, Z. & Campos, D. A. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature 437, 875–879 (2005).
- 25.
Wang, X., Campos, D. A., Zhou, Z. & Kellner, A. W. A. A primitive istiodactylid pterosaur (Pterodactyloidea) from the Jiufotang Formation (Early Cretaceous), northeast China. Zootaxa 1813, 1–18 (2008).
- 26.
Wang, X. & Lü, J. Discovery of a pterodactylid pterosaur from the Yixian Formation of western Liaoning, China. Chinese Sci. Bull. 46, 1–6 (2001).
- 27.
Rodrigues, T., Jiang, S., Cheng, X., Wang, X. & Kellner, A. W. A. A new toothed pteranodontoid (Pterosauria, Pterodactyloidea) from the Jiufotang Formation (Lower Cretaceous, Aptian) of China and comments on Liaoningopterus gui Wang and Zhou, 2003. Hist. Biol. 27(6), 782–795 (2015).
- 28.
Lü, J., Xu, L. & Ji, Q. Restudy of Liaoxipterus (Istiodactylidae: Pterosauria), with comments on the Chinese istiodactylids pterosaurs. Zitteliana B28, 229–241 (2008).
- 29.
Rodrigues, T. & Kellner, A. W. A. Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys 308, 1–112 (2013).
- 30.
Wellnhofer, P. The illustrated encyclopedia of pterosaurs. London: Salamander Books. 192 pp. (1991).
- 31.
Unwin, D. M., Frey, E., Martill, D. M., Clarke, J. B. & Riess, J. On the nature of the pteroid in pterosaurs. Proc. Royal Soc.London B 263, 45–52 (1996).
- 32.
Bennett, S. C. Articulation and Function of the Pteroid Bone of Pterosaurs. J. Vert. Paleontol. 27, 881–891 (2007).
- 33.
Peters, D. A reinterpretation of pteroid articulation in pterosaurs. J. Vert. Paleontol. 29, 1327–1330 (2009).
- 34.
Holgado, B. et al. On a new crested pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria. Sci. Rep. 9, 4940 (2019).
- 35.
Kellner, A. W. A., Weinschütz, L. C., Holgado, B., Bantim, R. A. & Sayão, J. M. A new toothless pterosaur (Pterodactyloidea) from Southern Brazil with insights into the paleoecology of a Cretaceous desert. An. Acad. Bras. Ciênc. 91, e20190768 (2019).
- 36.
Young, C. C. On a new pterosaurian from Sinkiang, China. Vertebrat. PalAsiatic. 8, 221–256 (1964).
- 37.
Bestwick, J., Unwin, D. M., Butler, R. J., Henderson, D. M. & Purnell, M. A. Pterosaur dietary hypotheses: a review of ideas and approaches. Biol. Rev. 93(4), 2021–2048 (2018).
- 38.
Dalla Vecchia, F. M. Cearadactylus? ligabuei nov. sp., a new early Cretaceous (Aptian) pterosaur from Chapada do Araripe (Northeastern Brazil). B. Soc. Paleontol. Ital. 32, 401–409 (1993).
- 39.
Amiot, R. et al. Oxygen and carbon isotope compositions of middle Cretaceous vertebrates from North Africa and Brazil: ecological and environmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 297, 439–451 (2010).
- 40.
Wang, X., Kellner, A. W. A., Jiang, S. & Cheng, X. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Sci. Nat. 99, 249–257 (2012).
- 41.
Wang, X., Rodrigues, T., Jiang, S., Cheng, X. & Kellner, A. W. A. An Early Cretaceous pterosaur with an unusual mandibular crest from China and a potential novel feeding strategy. Sci. Rep. 4, 6329 (2014).
- 42.
Frey, E., Buchy, M. C., Stinnesbeck, W., González, A. G. & Stefano, A. Muzquizopteryx coahuilensis n.g., n.sp., a nyctosaurid pterosaur with soft tissue preservation from the Coniacian (Late Cretaceous) of northeast Mexico (Coahuila). Oryctos 6, 19–40 (2006).
- 43.
Wang, X. & Zhou, Z. Pterosaur assemblages of the Jehol Biota and their implication for the Early Cretaceous pterosaur radiation. Geol. J. 41, 405–418 (2006).
- 44.
Kellner, A. W. A. & Campos, D. A. The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297, 389–392 (2002).
- 45.
Wellnhofer, P. & Kellner, A. W. A. The skull of Tapejara wellnhoferi Kellner (Reptilia, Pterosauria) from the Lower Cretaceous Santana Formation of the Araripe Basin, Northeastern Brazil. Mitt. Bayer. Staatssamml. Paläontol. Hist. Geol. 31, 89–106 (1991).
- 46.
Vullo, R. et al. A New Crested Pterosaur from the Early Cretaceous of Spain: The First European Tapejarid (Pterodactyloidea: Azhdarchoidea). PLoS One 7, e38900 (2012).
- 47.
Wang, X., Kellner, A. W. A., Zhou, Z. & Campos, D. A. Discovery of a rare arboreal forest-dwelling flying reptile (Pterosauria, Pterodactyloidea) from China. Proc. Natl. Acad. Sci. USA 105, 1983–1987 (2008).
- 48.
Kellner, A. W. A. & Calvo, J. O. New azhdarchoid pterosaur (Pterosauria, Pterodactyloidea) with an unusual lower jaw from the Portezuelo Formation (Upper Cretaceous), Neuquén Group, Patagonia, Argentina. An. Acad. Bras. Ciênc. 89, 2003–2012 (2017).
- 49.
Witton, M. P. & Naish, D. A. Reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS One 3, e2271 (2008).
- 50.
Edmund, A. G. Dentition. In: Biology of the Reptilia (eds Gans, C., Bellairs, A. A. & Parsons, N.), 117–200 (Academic Press, London, 1969).
- 51.
Bramble, D. M. & Wake, D. B.. Feeding mechanisms of Lower Tetrapods. In: Functional Vertebrate Morphology (eds Hildebrand, M., Bramble, D. M., Liem, K. F. & Blake, D. B.), 230–261 (Belknap, Harvard, 1985).
- 52.
Hiiemae, K. M. & Crompton, A. W. Mastication, food transport and swallowing. In: Functional Vertebrate Morphology (eds Hildebrand, M., Bramble, D. M., Liem, K. F. & Blake, D. B.), 262–290 (Belknap, Harvard, 1985).
- 53.
Evans, A. R. & Sanson, G. D. The effect of tooth shape on the breakdown of insects. J. Zool. 246, 391–400 (1998).
- 54.
Bennett, S. C. A second specimen of the pterosaur Anurognathus ammoni. Paläont. Z. 81, 376–398 (2007).
- 55.
Wilson, D. S. The adequacy of body size as a niche difference. Am. Nat. 109(970), 769–784 (1975).
- 56.
Barclay, R. M. R. & Brigham, R. M. Prey detection, dietary niche breadth, and body size in bats: why are aerial insectivorous bats so small? Am. Nat. 137(5), 693–703 (1991).
- 57.
Dial, K. P. Evolution of avian locomotion: correlates of flight style, locomotor modules, nesting biology, body size, development, and the origin of flapping flight. Auk 120(4), 941–952 (2003).
- 58.
Witton, M. P. & Habib, M. B. On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness. PLoS One 5, e13982 (2010).
- 59.
Xavier, J. C., Croxall, J. P., Trathan, P. N. & Wood, A. G. Feeding strategies and diets of breeding grey-headed and wandering albatrosses at South Georgia. Marine Biology 143, 221–232 (2003).
- 60.
Sustaita, D., Rico-Guevara, A. & Hertel, F. Foraging Behavior. In: Ornithology: Foundation, Analysis, and Application (eds Morrison M. L., Rodewald A. D., Voelker G., Prather J. F. & M. R. Colón), 439–492 (Johns Hopkins University Press, 2018).
- 61.
Goulding, M. & Ferreira, E. J. G. Shrimp-eating fishes and a case of prey-switching in Amazon Rivers. Rev. Bras. Zool. 2(3), 85–97 (1984).
- 62.
Bergmann, G. T. Trophic ecomorphology of the nonindigenous Mayan cichlid ‘Cichlasoma (Nandopsis)’ urophthalmus (Gunther 1862) in southern Florida. M.S. Thesis. University of South Florida, Tampa, Florida, 171 pp. (2002).
- 63.
Ross, A. J., Jarzembowski, E. A. & Brooks, S. J. The Cretaceous and Cenozoic record of insects (Hexapoda) with regard to global change. In: Biotic response to global change: the last 145 million years (eds Culver, S. J. & Rawson, P.F.), 288–302 (Cambridge University Press, 2000).
- 64.
Labandeira, C. C. The Fossil Record of Insect Extinction: New Approaches and Future Directions. Am. Entomol. 51(1), 14–29 (2005).
- 65.
Dalla Vecchia, F. M., Venturini, S. & Tentor, M. The Cenomanian (Late Cretaceous) Konservat-Lagerstätte of En Nammoûra (Kesrouane Province), northern Lebanon. B. Soc. Paleontol. Ital. 41(1), 51–68 (2002).
- 66.
Garassino, A. The macruran decapod crustaceans of the Upper Cretaceous of Lebanon. Paleontologia Lombarda 3, 1–40 (1994).
- 67.
Audo, D. & Charbonnier, S. New nisto of slipper lobster (Decapoda: Scyllaridae) from the Hadjoula Lagerstätte (Late Cretaceous, Lebanon). J. Crustac. Biol. 32(4), 583–590 (2012).
- 68.
Kushlan, J. A. Feeding ecology of wading birds. In: Wading Birds (ed. Kushlan, J.), 249–297 (National Audubon Society, New York, 1978).
- 69.
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32(3), 221–238 (2016).
- 70.
Philip, J. & Floquet, M. Late Cenomanian (94.7–93.5) In: Atlas Peri-Tethys Palaeogeographical Maps (eds Dercourt, J. et al.), 129–136 (Ccgm/Cgmw, Paris, 2000).
- 71.
Cohen, K. M., Finney, S. M., Gibbard, P. L. & Fan, J. X. The ICS International Chronostratigraphic Chart. Episodes 36, 199–204 (2013).
Acknowledgements
We would like to acknowledge L. Allan Lindoe for the artful and painstaking preparation of the specimen and Toni Farraj (Misopoint) for the photos that illustrate this paper. Special thanks to Hugo Salais-López (Metazoa Studio) for the outlines used in the figures. We thank the specimen acquisition fund, supported by donations from the Annual Palaeontological Gala at the University of Alberta, for assistance with preparation costs for the specimen. B.H. would especially like to acknowledge Rodrigo V. Pêgas (Universidade Federal do ABC) due to the discussions about pterosaur feeding behavior and lifestyle, and Julius T. Csotonyi, for the reconstruction of the new species (Fig. 5). The Fundação Carlos Chagas Filho de Amparo à Pesquisa do Rio de Janeiro (FAPERJ no. E-26/202.905/2018 to A.W.A.K.) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq #420687/2016-5 and #313461/2018-0 to A.W.A.K., #140789/2016-2 to B.H., and #311715/2017-6 to J.M.S.) partially funded this project.
Source:
Scientific Reports volume 9, Article number: 17875 (2019)





{kind=link}